

Краткое описание строения и свойств ферментов
Ферменты, или энзимы — это чаще всего молекулы белковой природы, катализирующие химические реакции в живых системах. Кроме белков, существует немногочисленная группа РНК ферментов.
Как и все белки, ферменты синтезируются изначально в виде полипептидной цепочки — первичная структура. Потом за счёт образования водородных связей и дисульфидных мостиков между между молекулами аминокислот в цепи образуется вторичная структура фермента. В дальнейшем под действием Ван-Вер-Вальсовых, электростатических и нековалентных сил образуется третичная и четвертичная структуры ферментов.
В каталитическом процессе участвует не весь фермент, а только его малая часть — активный центр.
Кроме этого стоит понимать, что любой фермент обладает высокой специфичностью и активностью. Для того чтобы катализировать реакцию, фермент связывается только с одним или несколькими определёнными субстратами. Большинство ферментативных процессов идёт со скоростями в 108-1020 раз большими, чем если бы они протекали сами. Говоря об активности ферментов, то их эффективность как катализаторов зачастую выше в 108-109 раз, чем их неорганические аналоги.
Кинетика ферментативных процессов
В основе кинетического анализа ферментативного процесса лежит предположение о том, что между ферментом (E) и субстратом (S) происходит обратимое образование фермент-субстратного комплекса. В дальнейшем в результате протекания реакции субстрат необратимо преобразуется в продукт (P) и комплекс распадается.
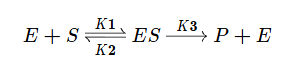
Используя допущение о стационарном протекании реакции, применим метод квазистационарных концентраций для фермент-субстратного комплекса.
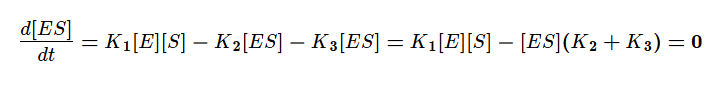
Учтём тот факт, что фермент при реакции находится как в свободной форме, так и в форме фермент-субстратного комплекса. Выразим тогда его свободную форму и подставим в квазистационарное уравнение.
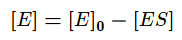
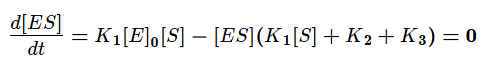
Выразим из этого уравнения [ES].
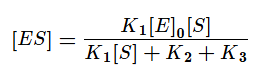
Скорость ферментативной реакции в основном определяется скоростью образования продукта реакции из субстрат ферментного комплекса. Тогда:
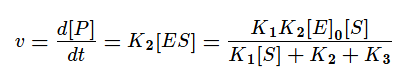
Разделим в получившемся уравнении и числитель, и знаменатель на K1 и получим:
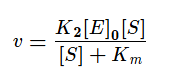
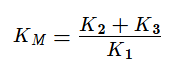
KM — это константа Михаэлиса, показывающая концентрацию субстрата, при которой скорость реакции равна половине максимальной. Численное значение KM зависит от многих параметров:
- pH среды, где протекает реакции;
- Температуры, при которой проводится реакция;
- Концентрации реагента, который подвергается ферментативному преобразованию;
- Концентрация продукта ферментирования;
- Наличию или отсутствию активаторов/ингибиторов ферментов
В результате действиях всех этих факторов KM может меняться в приделах от 1 до 10-8 моль*л-1. Стоит также отметить, что чем меньше значение KM, тем больше сродство фермента к субстрату, и наоборот. Кроме этого, ферментативные реакции часто проводят в буферных растворах с контролируемой ионной силой, чтобы избежать изменения KM (это можно прочитать в конспекте Гусаровой А.Р. «Расчёт осмотического давления и других коллигативных свойств растворов»).
На начальных этапах, когда количество субстрата превышает количество фермента, а концентрация продукта стремиться к нулю, уменьшением концентрации субстрата можно пренебречь. Тогда:
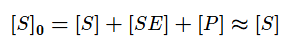
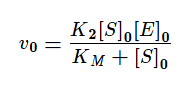
Если же K2>>K3, то в начале реакции устанавливается квазистатическое равновесие и константа Михаэлиса заменяется на субстратную константу (KS).
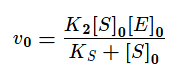
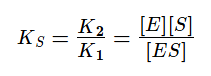
Проанализируем уравнение для v0 с KM для двух предельных случаев:
- [S]0 << KM, тогда реакция имеет первый порядок как по ферменту, так и по субстрату;

- [S]0 >> KM, в этом случае начальная скорость реакции не зависит от концентрации субстрата и называется максимальной скоростью ферментативной реакции vmax .Этот эффект так называемого субстратного насыщения обусловлен практически полным связыванием всего имеющегося в системе фермента в фермент-субстратный комплекс, поэтому его концентрация, а, следовательно, и наблюдаемая скорость реакции перестают зависеть от концентрации субстрата. Тогда

Это уравнение было получено Бриггсом и Холдейном в 1925 г., но названо ими уравнением Михаэлиса−Ментена. На практике же, невозможно никогда достичь vmax, так как работа фермента очень сильно зависит и от концентрации продукта реакции. Чем больше концентрация продукта, тем меньше сродство фермента к субстрату. Следует также помнить, что начальная скорость ферментативной реакции часто определяется по оптической плотности (это можно прочитать в конспекте Борисова В.В. «Построение градуировочного графика в спектрофотометрии и его статистическая обработка»).
На рисунке представлена зависимость начальной скорости ферментативной реакции от начальной концентрации субстрата.
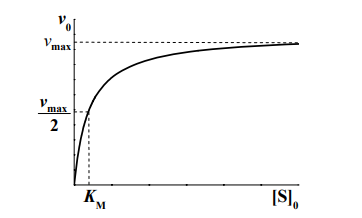
Стоит обратить внимание, что построение такого графика для определения vmax и KM на практике очень сложно. Поэтому для определения KM и vmax графически используют преобразованное уравнение Михаэлиса-Ментена, предложенное Г.Лайнуивером и Д.Бэрком для линеаризации графика.
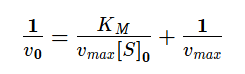
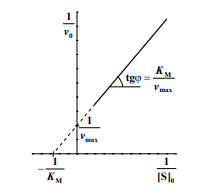
Графическое определение KM и vmax в координатах Лайнуивера-Берка
Максимальную скорость можно определить по отрезку, отсекаемому на оси ординат, 1/vmax , а константу Михаэлиса можно вычислить либо по тангенсу угла наклона прямой, равному KM/vmax , либо по отрицательному отрезку, отсекаемому на оси абсцисс.
Недостатки модели Михаэлиса
Несмотря на способность модели Михаэлиса-Ментена к упрощению анализа ферментативных процессов, её применение сопряжено с рядом методологических и практических ограничений.
- Уравнение можно применять только при стационарных условиях;
- Недопущение наличия продуктов ферментативной реакции в начальной системе;
- Не подходит для определения скорости реакции в реакциях с аллостерическими ферментами.
Заключение и выводы
- Ферменты — это сложные белковые молекулы, представляющие из себя биокатализаторы;
- Константа Михаэлиса — это константа, показывающая концентрацию субстрата, при которой скорость реакции равна половине максимальной;
- Уравнением Михаэлиса−Ментена имеет очень много допущений, вследствие чего теоретические значения, полученные при расчёте, могут сильно отличаться от полученных экспериментально;
- График получаемый при решении уравнения Михаэлиса−Ментена очень сложен для определения KM и vmax на практике, поэтому используют линеаризированное уравнение, предложенное Г.Лайнуивером и Д.Бэрком.
Пример решения задачи на уравнение Михаэлиса-Ментена
Фермент катализирует превращение субстрата S в продукт P по кинетике Михаэлиса–Ментена. При Vmax = 120 нмоль/мин и KM = 30 мкМ найдите начальную скорость реакции при концентрациях субстрата:
- [S]0 = 3 мкМ;
- [S]0 = 30 мкМ;
- [S]0 = 300 мкМ;
Решение задачи
Воспользуемся уравнением Михаэлиса-Ментена и подставим известные нам величины:
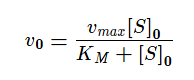
- V01=120*3/(30 + 3) = 360 / 33 ≈ 10.91 нмоль/мин;
- V02=120 · 30 / (30 + 30) = 3600 / 60 = 60 нмоль/мин;
- V03=120 · 300 / (30 + 300) = 36000 / 330 ≈ 109.09 нмоль/мин;
Комментарий
Следует помнить, что при [S]0 ≪ Km начальная скорость пропорциональна [S]; при [S]0 = Km начальная скорость равна половине Vmax; при [S]0 ≫ Km начальная скорость стремится к Vmax.